

Introduction
Materials and Methods
토양
작물
Results and Discussion
콩 생육 변화 분석
콩잎의 면적, 광합성 특성
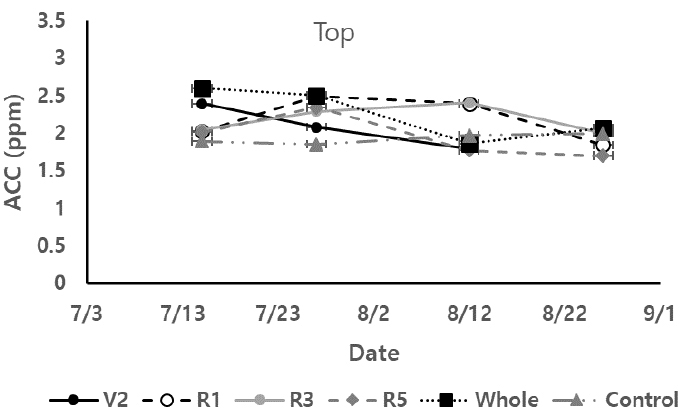
콩잎의 ACC 함량 특성
Conclusions
Introduction
기후변화로 인한 강우패턴의 변화는 전 세계적으로 나타나고 있는 현상이다. 국립기상과학원이 1912년부터 2017년 동안 우리나라 기후패턴을 비교한 결과 평균기온은 1.4°C 상승하였고 강수량은 20세기 초보다 124 mm 증가하였고 변동상이 커졌다. 강수량 변동성의 증가는 강한 강수는 증가하고 약한 강수는 감소하는 추세를 보이고 있다 (KMA, 2020). 이에 따라 작물재배 기간 동안 폭우로 인한 작물 생육과 수량 감소는 확대될 것으로 예상된다.
우리나라는 전체 농경지 면적 중 50% 이상은 논토양으로 이루어져 있고 담수로 벼를 재배하는 농사방법으로 인하여 토양배수가 불량하고 지하수위가 높은 특성을 갖고 있다. 이러한 토양은 적은 강수량에도 작물 근권부는 쉽게 과습조건이 되고 이는 뿌리의 산소공급이 부족하게 되어 작물에 과습 스트레스를 주게 된다 (Hiler and Clark, 1971; Belford, 1981; Chun et al., 2021a, 2021b). 지난 10년 동안 정부는 쌀의 과잉생산을 낮추고 밭작물의 자급률을 향상시키기 위해서 정책적으로 논에 밭작물을 재배하는 것을 지속적으로 장려하고 있다. 논에서 콩을 재배하는 면적은 매해 증가하여 2017년에 6.3천 ha, 2018년 8.9천 ha, 2019년에 11.5천 ha까지 증가하였다 (KOSIS, 2021). 하지만 논에서 작물들을 재배할 경우 위에 열거한 토양과 기상 조건들로 인하여 과습 스트레스가 쉽게 발생하고, 이로 인하여 작물의 생육 및 수량 감소가 발생한다 (Lone and Warsi, 2009; Palta et al., 2010; Ren et al., 2014; Ok et al., 2021; Sathi et al., 2022). Jung et al. (2011)은 배수불량 논에서 콩을 재배할 때 과습 스트레스 반응을 분석하였고 그 결과 콩은 논에서 토양수분이 증가함에 따라 생육과 수량 감소가 나타났고, 이를 바탕으로 콩이 수분 스트레스에 민감한 반응을 보인다고 결론지었다. Ahmed et al. (2015)과 Rhine et al. (2010)은 녹두와 콩을 과습조건에서 재배할 경우 수량이 20 - 39%까지 감소한다는 연구결과를 발표하였고, Board (2008), Buttery et al. (1993), Puiatti and Sodek (1999)은 과습으로 인한 콩 식물체내 N, P, K등 양분 함량이 감소한다고 밝혔다.
과습으로 인해 작물의 생육 및 수량의 감소는 작물 생육시기별로 다르다 (Umaharan et al., 1997). 생육시기별 과습 또는 침수 스트레스에 대한 반응은 다양하게 나타난다. 기존 연구에서는 유묘기와 생장기의 콩은 과습으로 인하여 잎면적, 건물중 그리고 수량 감소가 나타났고, 생육부진이 발생한다고 밝혔다 (Griffin and Saxton, 1988; Scott et al., 1989; Linkemer et al., 1998). 이러한 과습에 따른 작물의 생리적 반응과 수량감소는 주로 광합성 작용의 저하로 인한 것이라고 알려져 왔다 (Olivella et al., 2000). 최근에는 토양 내 과습조건이 뿌리환경에 영향을 주고 이는 줄기와 잎에 까지 생리적으로 영향을 주고 있다 (Castonguay et al., 1993). Vartapetian and Jackson (1997)은 과습환경에서 식물체 내에 생화학적 신호를 1-Aminocyclopropane-1-carboxilic acid (ACC)를 생성하여 뿌리에서 줄기, 잎으로 전달한다고 밝혔다. 토마토를 과습상태에 노출시키면 ACC가 생성되어 줄기와 잎에 도달하여 ethylene으로 변환되고 이는 줄기가 가늘게 생장하게 하고 잎의 비정상적 생육을 야기시키는 것으로 알려져 있다 (Bradford and Yang, 1981; Banga et al., 1996). 여러 연구들은 ACC와 ethylene은 과습환경에서 식물체 생장과 광합성 작용에 영향을 주고, 환경여건과 식물 종류에 따라 식물체에 다양한 영향을 준다고 밝혔다 (Abeles et al., 1992). 이에 따라 작물의 과습에 대한 반응, 특히 ACC의 반응은 다양할 것으로 예상할 수 있다. Ahmed et al. (2006)은 녹두의 과습에 따른 식물체내 ACC 함량과 광합성의 관계를 생육초기와 개화기 두 시기에 연구하였다. 그러나 이 연구에서도 상세한 생육시기별 과습에 따른 잎의 생리적 반응과 ACC 변화에 대한 연구, 특히 콩에 대한 연구는 자세히 이루어지지 않았다.
작물의 과습 스트레스에 대한 반응은 잎의 변형, 고사, 수량감소와 같은 피해가 나타나야 판별할 수 있었다. 과습피해가 나타나기 전에 빠른 과습 스트레스 판별을 위해서 본 연구는 과습상태에서 콩의 생육, 광합성 작용뿐만 아니라 ACC 함량의 변화를 조사하였다. 생육시기별 과습 스트레스에 대한 형태적, 생리적 변화를 분석하여 상관관계를 통해 과습 스트레스 판정을 위한 요인을 밝히고자 하였다.
Materials and Methods
토양
본 연구는 국립식량과학원 남부작물부 내 (경상남도 밀양시) 유리온실 내에서 2021년 5월부터 10월까지 수행하였다. 온실 내에 1 m (가로) × 1 m (세로) × 1 m (높이) 셀 안에 토양을 동일하게 채워 넣어 총 30개의 셀에서 콩을 재배하였다. 각 셀은 토양수분 센서 (Gropoint profile, ESS, Australia)를 설치하여 10, 20, 30, 40 cm 깊이에서 30분마다 토양수분 함량을 측정하였다. 30개 셀들은 개별적으로 토양수분 함량을 수분센서를 기반으로 자동으로 조절 및 제어가 가능하도록 자동관수시스템을 설치하였다. 이를 통해서 Table 1과 같이 생육시기별로 원하는 토양수분 함량이 유지되도록 하였다. 콩 파종 전에 토양시료를 채취하여 토성, pH, EC, 유기물, 유효인산 (available P2O5), 양이온 (K, Ca, Mg, Na)을 분석하였다. 토양분석은 농촌진흥청 토양화학 분석법 (NAAS, 2010)에 따라 검정하였다.
Table 1.
Soil moisture contents for soybean growth and excessive soil moisture treatments at various growth stages: V2, R1, R3, R5, Whole (whole growing period) and Control (no excessive soil moisture treatment).
| Growth stage | Soil moisture (%, v/v) |
| V2 | 24 (before treatment) - 70 (after treatment) |
| R1 | |
| R3 | |
| R5 | |
| Whole | 70 |
| Control | 24 |
토양 화학성을 분석한 결과 실험에 사용된 토양은 작물 재배에 적합한 화학성을 보였다 (Table 2). EC는 제한된 cell에서 반복적으로 비료투입과 작물재배를 하여 염농도가 증가한 것으로 판단된다. 염을 정상범위로 낮추기 위하여 파종 전에 반복적으로 침수와 배수를 반복하여 EC값을 1 이하로 낮추어서 실험을 하였다. EC외 다른 화학성 값들은 콩 재배에 영향을 주지 않은 범위 내에서 나타났다. 토성은 식양질 양토로 일반 논토양의 배수불량 특성을 갖도록 하였다. 우리나라 논토양은 배수불량 또는 약간불량 등급에 해당하는 면적이 전체 논토양 면적의 62%가 넘는다 (Jeon et al., 2002). 이러한 배수불랑의 논토양과 과습상태를 실험에서 재현하기 위해서 식양질 양토로 콩을 재배하였다.
Table 2.
Results of soil chemical and physical analyses. T-N means soil total nitrogen, O.M. means organic matter, Avail. P2O5 means available phosphorus, and Exch. cations means exchangeable cations.
작물
콩 품종은 대풍 2호 (Daepoong 2)이었고 2021년 6월 10일에 파종하였다. 각 셀에 18개의 콩을 1주2본으로 30 cm 간격으로 심어서 재배하였다. 파종 전 각 셀마다 시비량은 콩 시비처방기준 (RDA, 2016)에 따라 N-P2O5-K2O = 3-3-3.4 kg 10a-1 전량 기비하였다. 파종 후 콩 재배는 농촌진흥청 표준재배법에 준하였다. 과습처리 시기는 제2복엽기 (V2), 개화기 (R1), 협형성기 (R3), 협비대기 (R5)에 처리하고 수확할 때까지 과습상태를 유지하였다. 파종 직후부터 수확 때까지 전 생육기간 동안 과습처리한 전생육 (whole)과 대조구 (control)로 과습처리하지 않은 콩을 재배하였다. 콩 파종 후 모든 셀들은 24% 토양수분 함량을 유지하였다. Park et al. (2014)은 29콩 품종의 물 소모량을 조사한 결과 평균적으로 콩이 정상생육을 할 때 공극률의 50 - 60%를 필요로 한다고 하였다. 이에 따라 과습처리하지 않은 셀들과 Control 셀들은 토양의 공극률의 60%인 토양수분 센서값 24% (V/V)로 콩을 재배하였다. Whole 처리한 셀들은 파종 직후부터 토양수분 70% (공극률 150%)를 유지하여 수확 때까지 과습처리를 하였다. 각 셀은 과습처리 생육시기에 다다르면 토양수분 함량을 70%까지 증가하여 과습처리를 하였다. 이러한 과습상태는 수확시기까지 이어졌다. 모든 토양수분 조절은 셀내 자동 관수시스템으로 인하여 자동으로 조정되고 일정 토양수분 함량을 유지하였다.
과습처리 일주일 후 콩잎의 광합성률 (photosynthetic rate), 기공전도도, 엽록소 형광반응량을 측정하였다. 처리별로 줄기에서 맨 하위, 중간 및 상부에 위치한 잎을 대상으로 잎의 생리적 특성을 측정하였다. 광합성률과 기공전도도는 휴대용 광합성 측정기 (LI-6800, LI-COR Biosciences, NE, USA)를 사용하여 측정하였고 엽록소 형광반응량은 iFL Integrated Fluorometer (ADC BioScientific Ltd., UK)을 이용하여 측정하였다. 측정 전 측정대상 잎에 sample clip으로 약 20분간 광을 차단한 후 측정할 때는 2,000 µmol m-2 s-1의 광을 조사하였으며, 광화학반응효율 (Fv/Fm)값을 측정하여 분석하였다 (Fm: 최대형광반응, Fv: 최소와 최대형광반응 차이). 광합성률, 기공전도도, 엽록소 형광반응량을 측정한 잎들은 -80°C로 냉동시킨 후 ACC 함량을 경북대학교에서 분석하였다. 얼린 식물체는 분쇄 후 80% methanol 5 mL로 균질화하였다. 원심분리기로 상등액을 축출하고 C18 분말을 상등액에 첨가하였다. 이후 vortexing으로 다시 상등액을 축출하고 상등액을 Spin-vacuum을 통해 건조시켰다. 건조된 형태의 시료에 H2O 200 µL, buffer (0.2 M borate buffer (pH 8.5)) 300 µL, 유도체화 시약 (1.0 mM fluorescamine) 360 µL을 넣고 HPPLC-FLD로 분석하였다, 콩은 수확 후 생육 및 수량구성요소를 조사하였다.
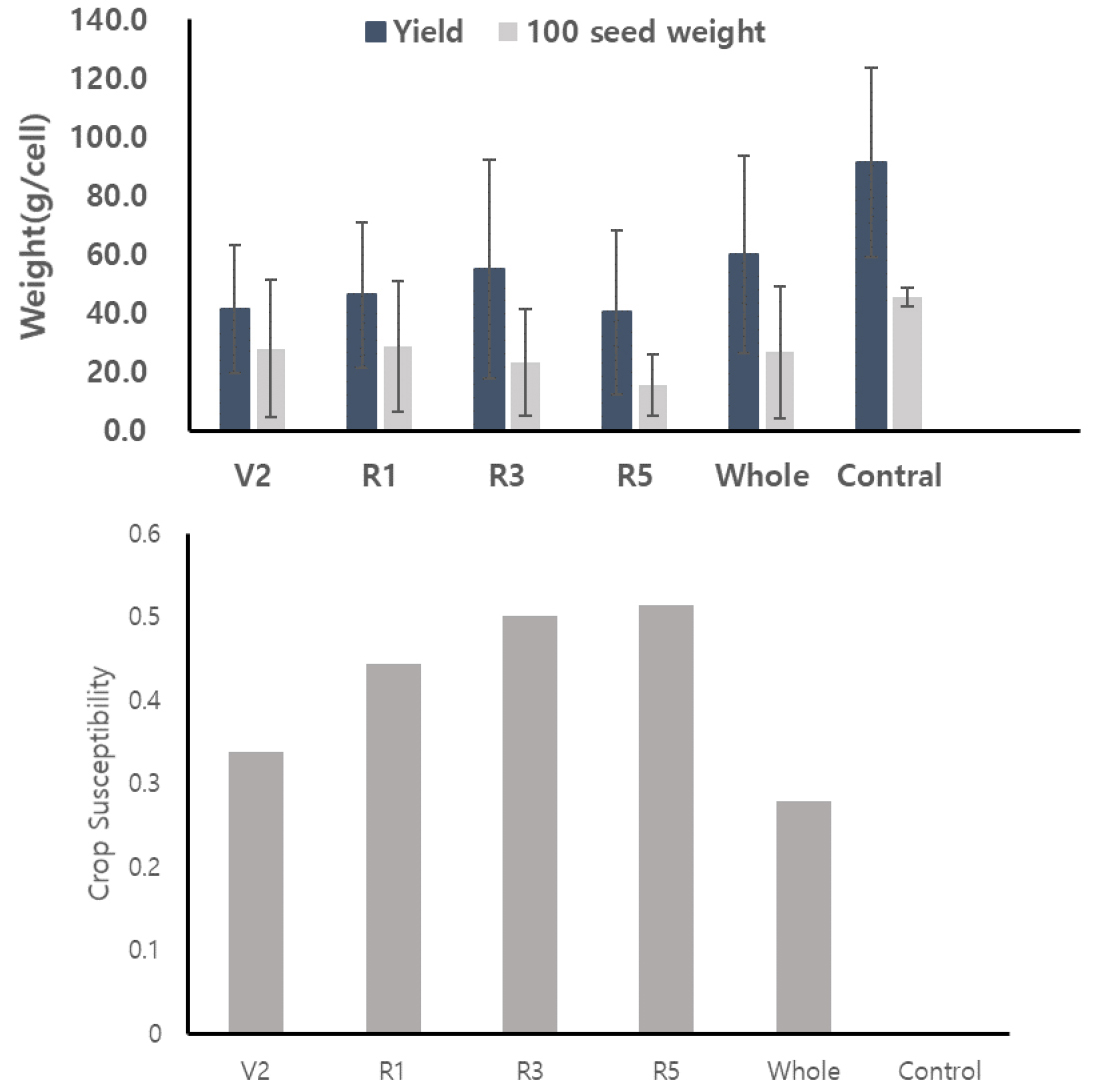
수분 스트레스에 의한 수량 감소율은 작물 민감성 요인 (crop susceptibility factor, CS)으로 계산하였다. 작물의 주요 생육기간 동안에 스트레스를 받은 작물의 수량을 조사하여 다음과 같이 계산하였다 (Hiler, 1969).
여기서, : 생육기에 임계 스트레스를 받은 작물의 수량
: 아무런 스트레스를 받지 않은 경우의 작물의 수량
통계 분석은 SPSS program (IBM, v.26, New York, USA)을 사용하여 ANOVA (analysis of variance)를 95% 유의수준에서 분석하였다.
Results and Discussion
콩 생육 변화 분석
콩 생육에서 경장은 과습처리를 받지 않은 Control, R3, R5에 과습 스트레스를 받은 콩들이 더 큰 값을 보였다 (Table 3). R3와 R5는 경장 성장이 다 이루어진 이후에 과습 스트레스를 받아 경장에는 영향을 주지 않은 것으로 판단된다. 반면 V2와 Whole 콩들의 경장값은 각 25.0 ± 16.33 cm, 27.7 ± 17.42 cm였다. 이러한 결과는 R3, R5, Control과 비교하여 확연히 작은 경장값을 보였다 (p < 0.05). 생육기에 과습 스트레스를 받을 경우 콩 경장과 같은 생육에 영향을 주는 것으로 나타났다. Shao et al. (2013)은 밀의 생육시기별 과습 스트레스 반응을 조사한 결과, 생육 초기와 개화기 전에 과습처리를 한 경우 뿌리 생육, 식물체 크기 등에서 확연한 감소가 나타났고, 개화기 이후 생식기간 동안 과습처리한 밀의 생육과 비교하여 더 큰 스트레스 반응을 나타낸 것을 밝혔다. 성장기의 과습은 뿌리의 성장과 호흡을 방해하고 이는 직접적으로 식물체의 성장에 영향을 주는 것으로 판단된다 (Palta et al., 2010). Linkemer et al. (1998)은 콩의 생육시기별 과습처리한 결과 성장기 (V2, V7)에 과습처리한 콩의 생육이 생식기 (R1 - R7)에 과습처리한 콩보다 경장이 더 작은 것으로 나타났다. 본 연구에서도 성장기에 과습처리한 콩들이 생식기에 과습처리받은 콩들보다 더 생육이 안 좋았고, 이는 낮은 경장으로 나타났다.
Table 3.
Growth characteristics of soybean with excessive soil moisture treatment at various growth stages; V2, R1, R3, R5, Whole (whole growing period) and Control (no excessive soil moisture treatment).
협수는 처리별로 차이가 나타나지는 않았으나 빈 협수는 R5에 가장 높게 나타났고 다음으로 R3, R1의 빈 협수가 높게 나타났다. 이와 더불어 고사율 또한 R5에서 65.9%로 가장 높게 나타났고, R3와 R1 시기가 다음으로 높은 고사율을 보였다. 이와 반대로 V2와 Whole 콩은 상대적으로 적게 고사 하였다. R1 이후 콩이 과습 스트레스를 받은 경우 7일 이내 고사가 발생한 반면 V2와 Whole 콩들은 고사를 하지 않고 과습 환경에 적응하여 꾸준히 생장하고 생식기를 거친 것으로 판단된다. Ara et al. (2015)은 다양한 콩 품종의 내습성을 실험한 결과, 시험대상의 콩 품종 모두에서 R1보다 R3, R5에 과습을 처리한 경우 협수가 정상생육한 콩보다 20% 이상 감소하는 것으로 나타났다. 이와 같은 협수의 감소는 수량감소로 이어졌다고 보았다. 본 연구에서는 협수는 차이가 나타나지 않았으나 종실이 없는 빈 협수의 증가는 수량에도 영향을 준 것으로 판단된다.
수량은 Control이 가장 컸으며 다음으로 V2, Whole 순으로 나타났다 (Fig. 1). 과습 스트레스에 대한 작물의 감수성을 정량화한 CS 값을 계산한 결과 (Eq. 1), 콩은 R5에서 CS값이 가장 높았고 또한 수량이 정상과 비교하여 51%의 감소를 보였다. 반면 과습에 가장 감수성이 낮은 것은 Whole 콩으로 수량감소는 Control과 비교하여 약 27%의 감소를 보였다. 과습처리 기간이 상대적으로 짧은 R3와 R5는 수량감소가 가장 컸고 과습 스트레스에 대한 감수성도 가장 높았다. Evans et al. (1990, 1991)은 콩과 옥수수의 생육시기별 과습 스트레스 반응을 정량화 하기 위해서 수량감소율에 따른 CS값을 계산한 결과, 옥수수는 V2 - V5에, 콩은 R5시기에 CS값이 가장 크며, 과습에 가장 민감한 반응을 보였다고 밝혔다. 백립중은 수량과 비슷한 경향을 보였다. 생육 후기에 과습이 처리되면 백립중 감소도 더 컸다 (Fig. 1). Control은 45.52 g으로 가장 큰 백립중을 보였다 (p < 0.05). R5의 백립중 값은 15.45 g으로 정상과 비교하여 65%의 감소가 나타났고, 이는 가장 작은 백립중값이었다. 반면 Whole, V2, R1은 상대적으로 백립중의 감소가 작았다. 수량과 백립중의 결과에서, 성장기에 과습처리 시 생육과 종실생산에 과습 스트레스 영향이 상대적으로 적은 것으로 판단된다. 이는 과습처리가 성장기에 이루어질 경우 콩이 과습환경에 적응하여 생육 및 종실 생산을 하는 것으로 판단된다. 기존 연구들에서 생육시기별로 과습처리 시 수량감소율은 개화기 이후에 더 크게 나타나고, 이는 생식기 동안 과습 스트레스에 더 민감하게 반응한다고 공통적으로 결론지었다 (Griffin and Saxton, 1988; Scott et al., 1989; Linkemer et al., 1998). 과습 스트레스를 받은 콩 종실의 크기는 수량과 같이 종실크기 또한 감소한다고 알려져왔다. Ara et al. (2015)은 R1보다 R3에 과습처리 시 수량과 백립중이 더 감소한다고 밝혔고 Sathi et al. (2022)은 12개의 콩 품종들을 과습처리한 결과 협수, 백립중, 수량이 모두 정상생육한 콩과 비교하여 감소하는 결과를 발표하였다. 본 연구의 생육시기별 과습처리에 따른 수량과 백립중 결과는 기존의 연구결과들과 같은 결과를 보였다.
콩잎의 면적, 광합성 특성
V2와 Whole 콩잎은 다른 콩잎들보다 잎면적이 더 작은 값을 보였으나, Control과 비교하여 잎 크기에서 차이는 없었다 (p > 0.05) (Fig. 2). 다른 생육기간별 과습처리에 따른 잎면적은 Control보다는 더 큰 면적값을 보였다 (p < 0.05). 기존 연구의 과습에 따른 잎면적 결과는 일반적으로 잎면적이 감소하는 것으로 나타났다. Linkemer et al. (1998)은 유묘기에 과습처리한 콩잎은 생식기에 과습처리한 콩잎보다 생장속도가 낮았고 이는 식물체의 낮은 성장속도로 나타났다고 보고하였다. Wang et al. (2017)은 개화기에 과습처리하여 경장과 잎면적 모두 확연한 감소를 보였다고 밝혔다. Guang et al. (2012)은 개화기 이후 과습처리한 목화잎의 경우 잎면적이 증가한다는 결론을 지었다. 이 연구에서 과습에 따른 잎면적의 변화는 수량 감소와 관련이 적고 과습에 노출되는 시기와 기간이 수량에 더 큰 영향을 준다고 밝혔다. 따라서 과습에 상대적으로 적게 노출된 개화기, 협형성기, 협비대기의 콩잎 크기가 더 큰 값을 보인 것으로 판단된다.
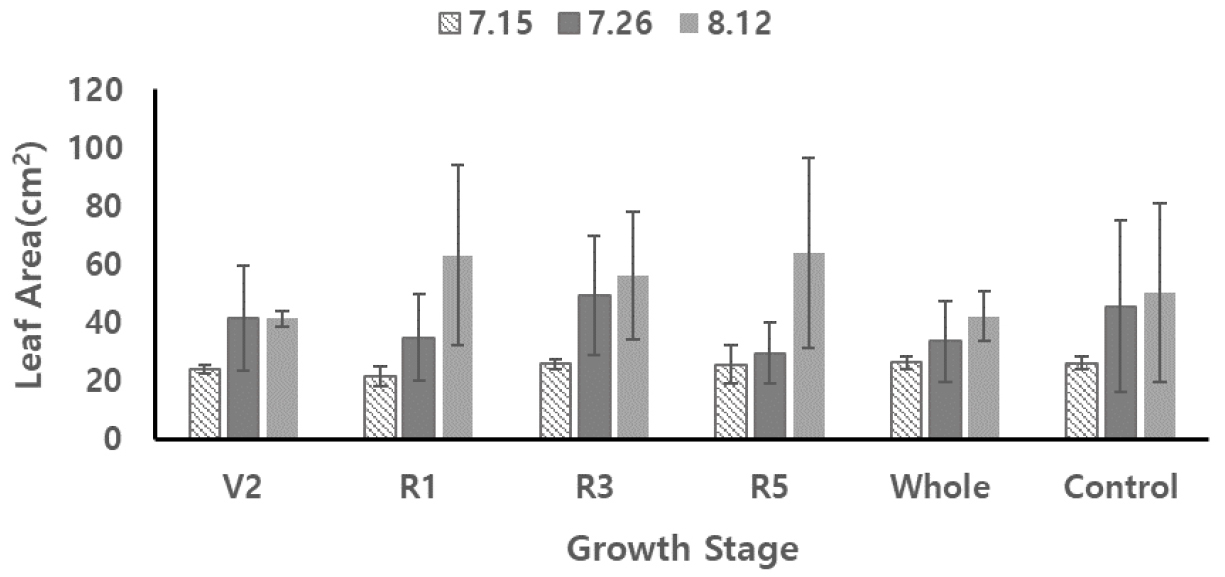
Fig. 2
Average and standard deviation results of soybean leaf area from excessive soil moisture treatments at five growth stages; V2, R1, R3, R5, whole growing period (whole) and control (no excessive soil moisture contents). Leaf areas were measured on July 15th (7.15), July 26th (7.26) and August 12th (8.12).
잎의 생리적 특성을 조사하고자 광합성률, 기공전도도, 엽록소형광 반응 (광화학반응효율, Fv/Fm)을 측정하였다 (Fig. 3). 파종 후 3주 후부터 측정한 결과 생육초기에는 모든 콩잎의 광합성률은 통계적 차이를 보이지 않고 20 µmol m-2 s-1 전후의 값들을 보였다. Whole과 V2에 과습처리 일주일 (7월15일) 후 광합성률이 각각 10.67 ± 6.51 µmol m-2 s-1, 9.57 ± 1.82 µmol m-2 s-1로 가장 낮았으나 이후 생육 중 후기에 이르러서는 증가하는 광합성률을 보였다. R3와 R5에는 과습처리 전까지는 Control과 비슷한 광합성률을 보였으나 과습처리 이후 (8월 12일, 8월 26일) 모두 급격한 광합성률 감소를 보였고 이와 같은 광합성률의 감소는 지속되었다. Control 콩은 생육초기와 중기까지 21 µmol m-2 s-1 전후 값을 유지하고 R1 이후 감소를 하여 R5에는 16.93 ± 2.73 µmol m-2 s-1 값을 보였다. Whole과 V2도 R5에는 Control과 비슷한 광합성률값을 보였으나 R3와 R5에 과습처리된 콩들은 광합성률값이 2.14 ± 2.07 µmol m-2 s-1, 2.80 ± 1. µmol m-2 s-1까지 낮아졌다. 기공전도도는 광합성률과 비슷한 경향을 보였으나 엽록소 형광 반응량은 과습처리에 따른 차이가 확연하게 나타나지 않았다 (p > 0.05).
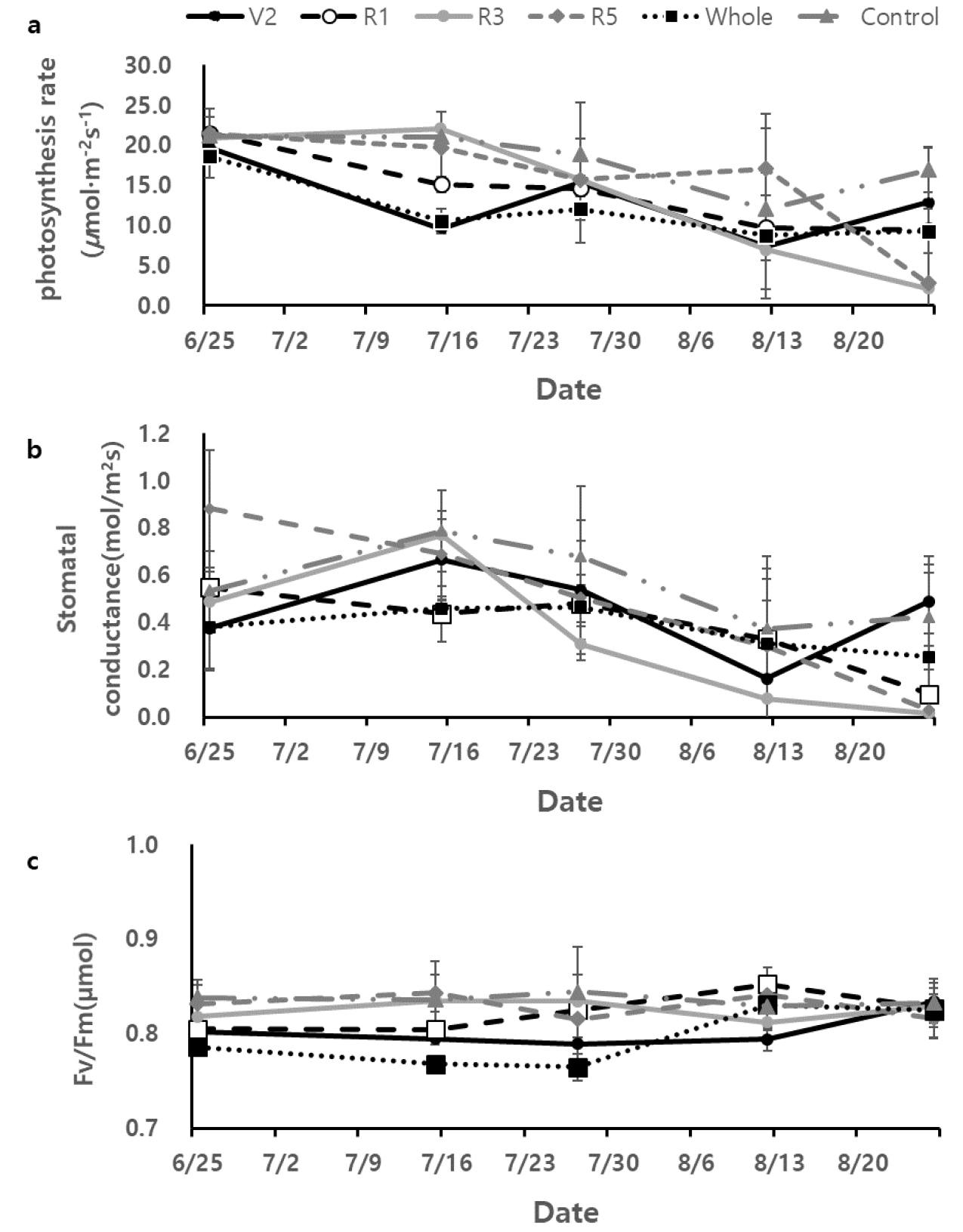
Fig. 3
Distributions of photosynthesis rate (a), stomatal conductance (b) and chlorophyll fluorescence (Fv/Fm, c) from soybean leaves with excessive soil moisture at five growth stages; V2, R1, R3, R5, whole growing period (whole) and control (no excessive soil moisture contents). Fm represents maximum variable fluorescence and Fv represents difference between maximum and minimum variable fluorescence.
콩잎의 ACC 함량 특성
ACC 함량을 과습처리 전후로 분석한 결과 과습처리 후 일주일 동안 ACC 값의 증가를 보인 후 ACC 값은 과습상태가 유지되면 2 ppm 전후 값으로 안정화가 되어갔다 (Fig. 4). 위치별로 잎의 ACC값은 상부쪽에 있는 잎보다 하부쪽으로 내려갈수록 과습에 따른 ACC값 변화가 더 컸다. R1에 과습처리한 콩의 경우 상부의 잎은 ACC값이 2.49 ppm이었고 중간잎은 2.68 ppm, 하부잎은 2.96 ppm이었다. 다른 생육시기에 처리한 콩잎도 하부의 잎이 과습에 따른 ACC 반응이 가장 컸다. 생육시기별 과습에 의한 잎의 ACC 반응은 다르게 나타났다. Control 콩은 전 생육기간 동안 평균 1.94 ± 0.21 ppm을 보였다. Control 콩은 ACC값이 평균값에서 크게 증가하거나 감소하는 값을 보이지 않았다. V2와 Whole 콩잎은 과습처리 일주일 후에 ACC값이 과습처리 하지 않은 ACC값보다 더 큰 값들을 보였다. 이와 같은 ACC 증가는 줄기의 상, 중, 하부 잎들에서 공통적으로 나타났다. 이후 생육이 진행되면서 과습조건에서도 ACC 값은 감소세를 나타냈고 R5시기 전후로 Control과 비슷한 ACC값을 보였다. R1와 R3는 과습처리 일주일 후 ACC값이 각각 2.96 ± 0.12, 2.73 ± 0.06 ppm으로 같은 시기 다른 콩잎들보다 높은 값을 보였다 (p < 0.05). R5에 과습처리된 콩잎은 과습처리 후에도 ACC값의 증가는 나타나지 않았다. 다만 R5에 과습처리된 콩잎은 과습처리 후 ACC값이 평균 1.47 ± 0.70 ppm 다른 처리의 콩잎들보다 더 낮게 나타났다 (p < 0.05). 이는 광합성, 엽록소 형광반응 분석과 같은 결과였다. Whole 콩잎에서는 과습처리 일주일 후 V2와 같이 ACC값이 2.35 ± 0.86 ppm으로 다른 처리의 콩잎들보다 가장 큰 ACC 값을 보였고, 이후 Whole 콩잎의 ACC값이 감소하여 Control과 비슷한 값을 보였다.
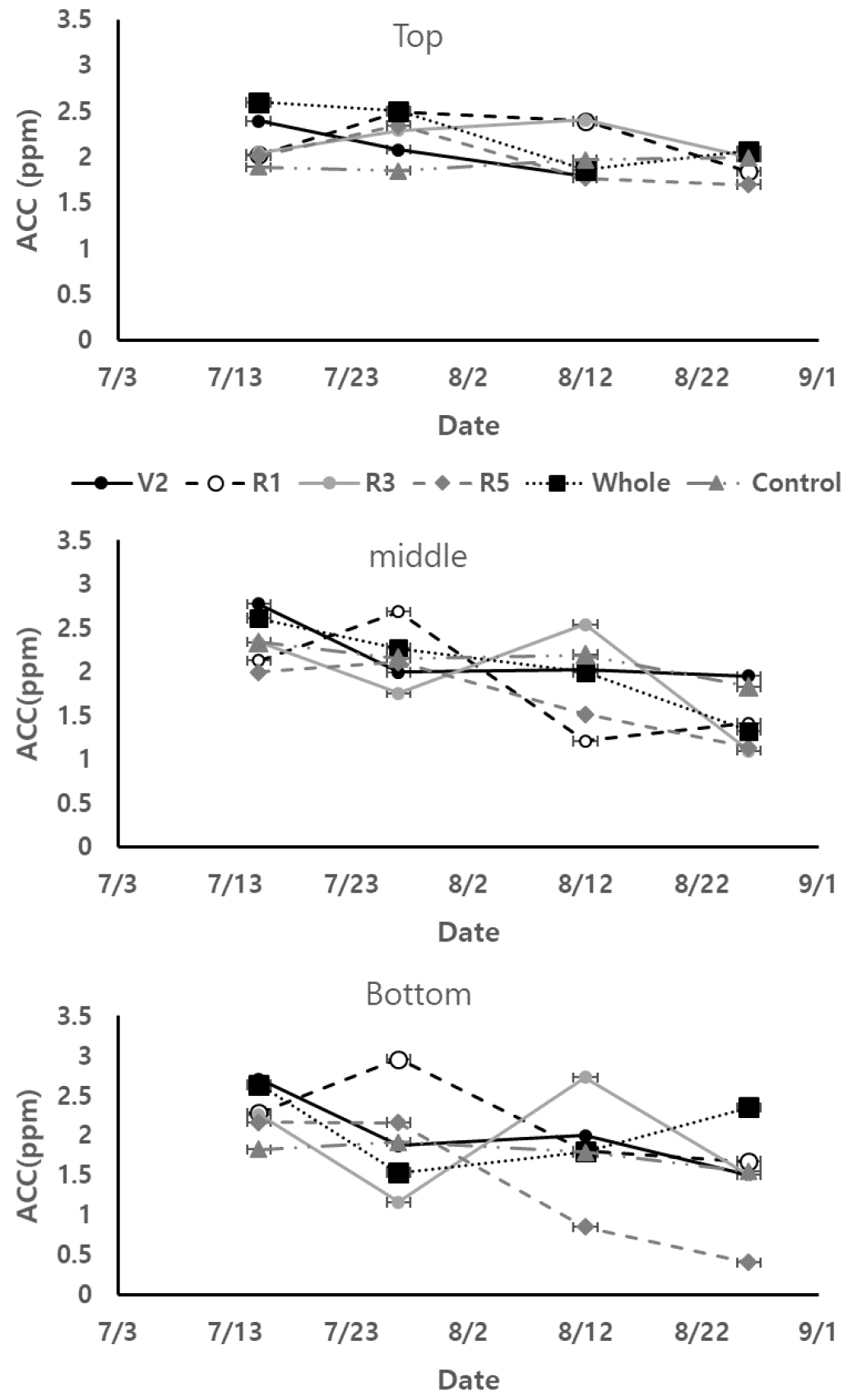
토양 내 과도한 토양수분은 콩잎의 광합성과 관련된 생리적 활동을 저하하는 것으로 나타났다. 광합성 활동이 저하가 되어짐과 동시에 ACC 함량은 과습으로 인한 증가를 보였다. 과습에 의한 잎의 생리적 변화는 14일 후 계속되는 과습상태에서도 광합성 활동량은 감소하고 ACC 함량은 증가하였다. Ahmed et al. (2006)은 과습처리 후 2일부터 녹두 잎의 광합성 활동이 저하되고 ACC 함량이 증가하고 이후 14일 동안 유지 또는 서서히 감소하여 16일째에 정상생육하는 녹두 잎과 같은 광합성 활동, ACC 함량이 감소한다고 연구결과를 밝혔다. 이러한 현상은 과습으로 인하여 뿌리 ACC 함량이 증가하고 이는 식물체 내의 광합성 활동량을 감소시키고 ACC 함량을 증가시키는 것으로 판단된다. Jackson (2002)은 ACC 이동은 과습상태의 식물체 잎에 추가 ethylene 형성을 돕고 이 ethylene은 뿌리의 스트레스 시그널을 식물체로 보내주는 역할을 한다고 밝힌바 있다. 따라서 과습을 받은 식물체는 뿌리의 ACC를 받아서 잎에 ACC를 축척하고 이 ACC는 공기와 만나서 ethylene을 형성하고 이는 결과적으로 광합성 활동량을 감소시킨 것으로 판단된다.
본 연구결과에서 과습처리에 따른 콩의 생육과 콩잎의 생리적 반응에서 Whole 콩과 R5에 과습처리된 콩의 반응은 다른 콩들과 다르게 보였다. V2, R1, R3에 과습처리된 콩잎은 과습 후 일주일에 광합성 활동량은 감소하고 ACC 증가를 보이고 14일 후 Control과 비슷한 광합성 활동량과 ACC 값을 보여주었다. 수량은 과습에 노출된 시기가 늦을수록 감소는 커졌다.
Whole 콩은 과습 초반에는 과습 스트레스 반응을 보이고 14일 후부터 콩잎의 생리적 반응은 Control과 비슷한 반응을 보였다. 줄기의 생육은 정상생육과 비교하여 낮았으나 다른 시기에 과습처리된 콩들보다 수량감소가 더 작았다. 기존 연구들에서 계속되는 과습환경은 식물체가 과습에 적응하는 형태를 보여준다고 밝혀왔다. 과습에 적응하기 위해 식물체는 잎의 생장, 줄기 생육, 뿌리의 통기구조 등을 변화시키는 것으로 알려져 있다 (Sathi et al., 2022). Voesenek and Bailey-Serres (2015)는 과습으로 인한 ethylene 생성, 식물체의 상위생장성, 백화현상 그리고 잎의 노화로 적응한다고 보고하였다. Luan et al. (2020)은 여러 품종의 콩이 과습에서 뿌리수를 더 증가하는 내습성을 보였고 ethylene 증가, abscisic acid 감소, 그리고 낮은 산소축적은 세포틈새 공간을 증가시키고 더 나은 엽록체 세포막 형성을 하게되어 과습 환경에 적응하게 한다고 결론지었다. 또한 Sairam et al. (2008)은 낮은 산화-환원 헤모글로빈과 산화질소 유기를 통해 과습 스트레스를 식물체가 더 잘 적응하도록 한다고 밝혔다. 콩이 생육 초반부터 장기간 과습에 노출된 경우 위와 같은 이유로 적응을 하게 되어, 줄기, 잎의 생장은 줄어드나 수량감소는 영향을 덜 받는 것으로 판단되다.
R5에 과습에 노출된 콩은 광합성 활동량이 감소하기 시작하여 수확 전까지 계속적인 감소를 보였다. ACC 함량 또한 과습에 노출된 이후 다른 시기에 과습처리된 콩들보다 더 낮은 ACC 함량을 나타냈다. 이러한 잎의 급격한 노화작용은 수량을 가장 크게 감소시키는 영향을 주었다. Rhine et al. (2010)은 5품종의 콩을 V5시기와 R5시기에 과습처리를 한 결과 V5 시기에 과습 받은 콩은 17%의 수량감소를 보였고, R5는 수량이 40%까지 감소한다고 밝혔다. VanToai et al. (2010) 또한 콩 21품종에서 R1 이후 과습처리한 콩들의 수량 감소가 생육 초기 과습처리한 콩들보다 더 컸다고 보고하였다. Sung (1993)은 R1과 R5시기에 과습처리한 콩잎에서 니트로게나아제 활동량을 측정한 결과 R1은 과습 10일처리후 니트로게나아제 활동량이 감소하였으나 과습처리를 제거하면 다시 정상적은 활동량으로 복원을 하였다. 반면 R5는 과습 4일만에 니트로게나아제 활동량이 매우 낮게 나타났고 토양수분을 정상으로 처리한 이후에도 니트로게나아제 활동량이 회복되지 못 한다고 보고하였다. 이는 R5시기에 과습처리를 받으면 식물체가 니트로게나아제 활동량의 감소와 질소 흡수가 막아지고 이는 식물체가 회복불능의 상태가 된다고 결론지었다. 본 연구에서도 R5에 과습처리된 콩의 고사율이 65.9%로 다른 시기에 과습처리된 콩보다 더 높은 고사율을 보였다. 이는 과습처리 후 잎의 광합성이 낮아지고 ACC 함량도 지속적으로 떨어지고 회복을 하지 못하여 고사에 이른 것으로 판단된다. 과습 스트레스를 받은 콩잎은 광합성 특성의 변화와 ACC 함량의 변화로 특성 지어질 수 있었다. 광합성 특성은 과습처리 차이에 따른 변화가 명확하게 나타나지 않았으나 ACC 함량 변화는 다른 특성들보다 명확한 차이를 보였다. 이에 따라 ACC 함량 변화를 과습으로 인한 스트레스 반응을 알려주는 지표로 활용 가능할 것으로 판단된다.
Conclusions
본 연구결과를 종합한 결과, 콩은 생육기간 중 R3와 R5 기간에 과습 스트레스를 받은 경우 수량감소가 가장 큰 것으로 나타났다. 전 생육기간 동안 과습 스트레스를 받으면 일시적으로 ACC 함량이 증가하고 광합성이 감소하는 특성을 보였다. R3, R5를 제외한 다른 생육기간에 과습을 받은 콩잎에서는 ACC와 광합성 활동이 스트레스를 받지 않은 콩잎의 상태로 되돌아가는 것으로 밝혀졌다. ACC 특성은 과습 스트레스를 받고 1주일까지 증가한 값을 콩잎의 생육시기들마다 공통적으로 나타났다. 이는 추후 콩의 과습 스트레스를 진단하는데 있어서 중요한 지표로 활용이 가능할 것으로 예상된다.
