

Introduction
Materials and Methods
수종 선정 및 육묘
양분집적 모델
묘목 측정 및 분석
통계분석
Results and Discussion
시비 처리에 따른 묘목 성장량 변화
시비 처리에 따른 묘목 내 양분함량 변화
Conclusions
Introduction
간척지는 원래 수도작을 위해 갯벌을 매립하였지만 최근 쌀 초과 공급에 의해 벼 재배보다는 밭 작물 재배로 전환하거나 산업, 관광, 도시로 개발 계획이 변경되고 있다 (Jeong et al., 2020; SDIA, 2023). 이와 더불어 산림청과 농식품부에서는 생태 ‧ 경관을 개선하고, 비산먼지 저감 및 풍해 예방을 위해 새만금 간척지에 2026년까지 228 ha 면적에 나무심기를 확대할 계획이며, 이는 농생명용지 묘묙장 (55 ha), 강 유역 방수제 도로사면 (73 ha), 농생명 용지 내부도로 (100 ha)를 포함한다 (KFS, 2021). 간척지 토양은 염농도가 높고, 지하수위가 높으며, 토양 구조 발달이 잘 되지 않아 배수가 불량하며 양분 함량이 낮아 밭 작물이나 나무가 자라기에는 적합하지 않다 (Jeong et al., 2020; Seo et al., 2023). 따라서 간척지 토양에 나무를 재배하기 위해서는 양분 공급을 위한 개량제 투입은 물론 잡초 제거를 위한 제초제 살포나 잡초매트 피복 등의 추가 비용이 필요하다.
양분집적 기술 (nutrient loading technique)은 묘목 육묘 단계에서 시기별 시비량 과 같은 시비 조건을 달리하여 식물체 생체량은 유사하지만 체내 양분 함량을 높여 양분 결핍 (deficiency) 상태의 묘목을 양분 잉여 (luxury consumption) 수준으로 육묘하는 기술이다 (Imo and Timmer, 2002). 관행적으로 재배된 묘목과 비교하면, 양분집적 기술로 재배된 묘목은 오래된 조직에 영양분을 보존하기보다는 당해연도 성장을 위해 영양분을 재이동하는 능력을 보인다 (Malik and Timmer, 1995, 1998). 육묘장에서 자란 묘목은 뿌리 시스템의 느린 재생과 확장으로 인해 토양 영양분의 이용과 흡수가 제한되기 때문에 포장 이식 직후 영양 결핍을 나타내는 경우가 많다 (Burdett et al., 1984; Burdett, 1990; Van den Driessche, 1991). 하지만, 양분이 집적된 묘목은 척박한 토양에 뿌리가 적응하는 동안 영양분의 재이동을 통해 집적된 양분을 이용함으로써 이식 후 생존율이나 성장량이 관행 육묘된 묘목에 비해 우수하다 (Malik and Timmer, 1995, 1998).
양분집적 기술은 묘목 육묘 시 시비량을 일정하게 유지하는 관행 방식과는 다르게 시비량을 생육 후기로 갈수록 지수함수로 늘려 줌으로써 (i.e., exponential model) 식물체내 양분을 집적할 수 있도록 한다 (Malik and Timmer, 1995, 1998). 하지만 지수함수 모형의 양분공급은 생육 초기 양분이 부족할 수 있고 생육 후기 급격한 시비량 증가로 인하여 식물체에게 독성을 유발할 수 있다. 이를 보완하기 위하여 제시된 모형 (i.e., modified exponential model)이 생육 초반 시비량을 일정 기간동안 보상한 후 시비량을 지수함수로 증가시키는 방식이다. 질소는 묘목을 포함한 식물 성장에 가장 중요하지만 결핍되기 쉬운 양분이다. 양분집적 기술을 이용한 높은 질소 시비율은 묘목 생장, 광합성율, 양분농도 및 뿌리 생장능력을 촉진할 수 있다 (Van den Driessche, 1988). 외국에서는 양분집적 기술을 활용하여 육묘된 넓은잎삼나무 (Chinese-fir, Cunninghamia lanceolata) (Xu and Timmer, 1999), 검은가문비나무 (black spruce, Picea mariana) (Malik and Timmer, 1998; Imo and Timmer, 2001; Salifu and Timmer, 2003), Lutz 가문비나무 (Lutz spruce, Picea glauca × lutzii) (Jonsdottir et al., 2013), 방크스소나무 (jack pine, Pinus banksiana) (Pokharel et al., 2017) 등 다양한 수종에 대한 포장 이식 후 생육 초기 성장에 대한 평가가 이루어졌다. 하지만, 우리나라에서는 관련 기술이 도입되지 않아, 대표적 고유 수종에 대한 양분집적에 대한 연구는 없다.
본 연구는 국내 대표 수종에 대한 유묘 양묘 과정 중 양분집적 가능 여부를 평가하기 위해 수행되었다. 양분 시비량을 일정하게 유지하는 관행 모델에 비해 유묘 성장 시기에 맞춰 양분 시비량을 증가시켜주는 지수 모델 또는 수정 지수 모델이 양분집적에 더 효율적일 것으로 가설을 세우고 연구를 진행하였다.
Materials and Methods
수종 선정 및 육묘
국내 대표 수종에 대한 양분집적 가능성 평가를 위하여 해송 (Pinus thunbergii), 상수리 (Quercus acutissima), 느티나무 (Zelkova serrata)를 선정하였다. 묘목은 충남 부여 소재 영림농원에서 포트 (43.2 × 26.7 × 10 cm, 구멍크기: 3 × 3 × 10 cm) 묘를 구매하였으며, 노지 이식 시기를 고려하여 해송은 1년생, 상수리와 느티나무는 발아 2주 유묘를 사용하였다. 양분집적 모델, 포장 이식 처리 및 육묘 중 고사율을 고려하여 수종 당 2,000묘를 육묘하였으며, 비닐하우스 내 온도, 습도, 일사량 등 불균질한 조건을 고려하여 묘목을 3등분하여 월 1회 위치를 서로 변경하였다. 성장량 측정 및 묘목 샘플 채취는 각 위치 (3 반복)에서 균등하게 진행하였다. 육묘는 전북대학교 농장 비닐하우스에서 2020년 6월부터 10월까지 진행하였다. 비닐하우스에 설치된 스프링클러와 분무기를 이용하여 기온을 고려하여6월 (주 3회), 7 - 8월 (1일 1회), 9 - 10월 (주 2 - 3회), 11월 이후 (월 1회)와 같이 다르게 관수하였다.
양분집적 모델
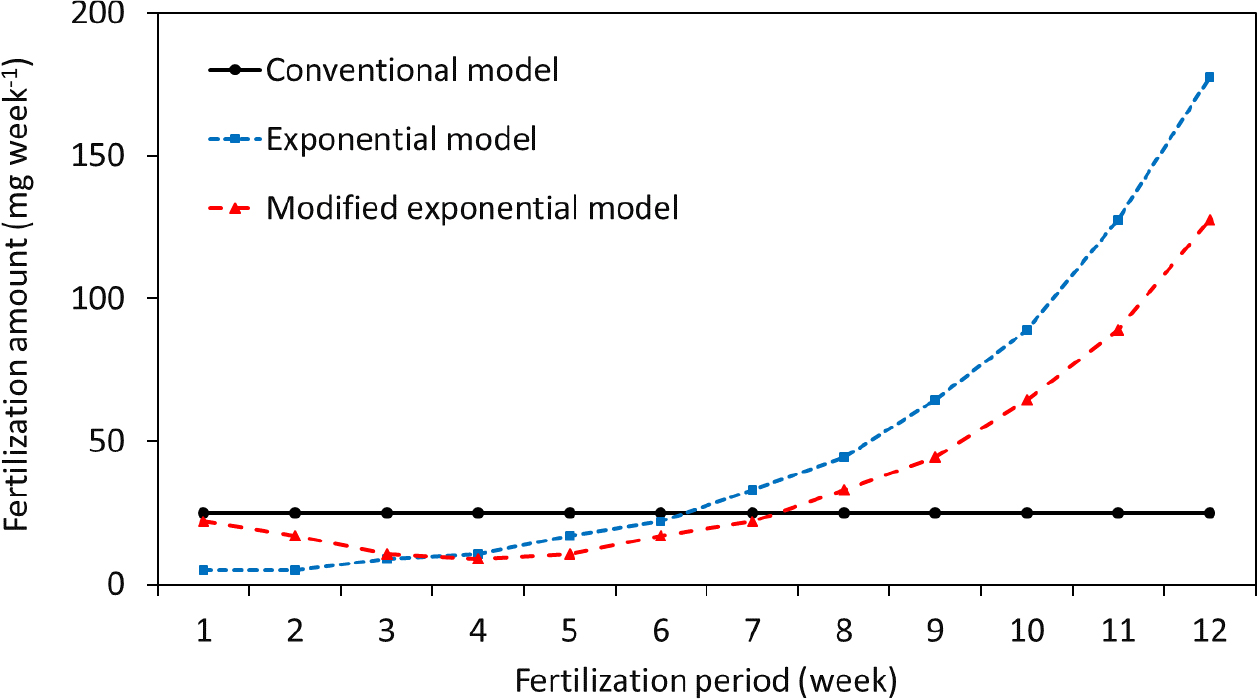
묘목 양분집적 가능성을 평가하기 위해 세 가지 시비 모델을 사용하였다. 육묘 기간 중 일정량을 나누어 시비하는 관행방식 (conventional fertilization, CF), 묘목 성장 시기에 맞춰 시비량을 지수함수꼴로 증가시키는 지수모형 (exponential fertilization, EF), 지수모형의 초기 시비량 부족에 의한 성장 저해를 보완한 수정지수모형 (modified exponential fertilization, MEF)을 사용하였다. 전통방식으로 육묘 시 투여되는 비료 양 (300 mg seedling-1, 육묘장 관행)을 고려하여 주차 별 시비량을 다음과 같이 계산하였다.
지수모형 (Timmer, 1997; Salifu and Timmer, 2003):
: desired amount to be added in a number (t) of fertilizer applications
: N content in seed or the initial N content, mg plant-1 season-1
: relative addition rate required to increase NS to a final N content (NT + NS)
: number of fertilization
본 연구에서는 관행 총 시비량 300 mg seedling-1, NS는 5 mg plant-1 season-1을 기준으로 하였으며, r은 0.34로 계산되었다.
각 시비 시기에 필요한 시비량:
: cumulative amount of N added up to the previous application
수정지수모형 (Timmer et al., 1991; Imo and Timmer, 1992; Jacobs and Timmer, 2005): 지수 모형과 동일하나 초기 보상기간 추가, 보상기간 계산식:
: amount of N called N compensation, initially subtracted from the last two applications calculated from Eq. 2
: final amount of nutrient added in the compensation period
: relative addition rate required to increase N0 to Nc
: compensation period
시비는 육묘장에서 많이 사용하는 크리스탈론 그린 (N18 + P18 + K18 + 미량원소, www.yara.kr)을 사용하였으며, 시비 모형 별 해당 양의 비료 (Fig. 1)를 증류수에 희석 후 디스펜서를 이용하여 5 mL씩 6월 첫째 주부터 매주 시비하였다. 모델별 육묘 기간 중 묘목 당 총 시비량은 관행, 지수, 수정 지수 모델에서 각각 300, 605, 467 mg seeding-1이었다 (Fig. 1).
묘목 측정 및 분석
처리별로 생육 시기에 따른 묘목 성장량을 평가하기 위하여 각 포트에서 1 - 2개 묘목을 무작위 선택 (수종별, 처리별 30개채)하여 묘묙 수고를 6월 2일, 6월 29일, 8월 3일, 9월 21일 눈금자를 이용하여 1 mm 단위까지 측정하였다.
시기별 묘목 내 양분 함량을 측정하기 위해 처리구의 수종 별로 각 5개 묘목을 수고 측정일에 채취하였으며, 채취 후 식물체를 흐르는 물과 증류수로 세척하여 먼지와 상토를 제거하였다. 식물체를 뿌리, 줄기, 잎 세 부분으로 분리한 후 오븐에서 60 °C로 무게가 일정해질 때까지 건조하였다. 건조된 시료의 건물중 측정 후 믹서기와 분쇄기 (MM400, Retsch, Germany)을 이용하여 분쇄하였다. 각 식물체 부위별 탄소 및 질소 농도를 원소분석기 (Flash EA 1112, Thermo Fisher Scientific Inc., Netherlands)로 분석하였으며, 탄소 및 질소 함량은 부위별 건물중 × 농도로 계산하였다.
통계분석
양분집적 모델에 따른 묘목 성장량 및 양분 함량 차이의 통계적 유의성은 분산분석 (ANOVA)으로 검증하였다. 통계분석은 IBM SPSS Statistics 27 프로그램을 이용하여 유의확률 95% 수준에서 검토하였다.
Results and Discussion
시비 처리에 따른 묘목 성장량 변화
시간에 따라 묘목 수고는 증가 (P < 0.001)하였다 (Fig. 2). 상수리와 느티나무의 성장량 증가가 해송에 비해 급격하게 나타났는데, 이는 해송은 1년 성장 묘목을 대상으로 양분집적 육묘를 하였지만, 상수리와 느티는 발아 2주 된 유묘를 사용하여 해송에 비해 급격하게 성장량이 증가한 것으로 판단된다.
해송과 상수리는 양분집적 모델에 따른 묘목 수고 차이가 없었으나 느티나무는 지수모델 시비 처리구에서 관행과 수정지수모델에 비해 12주차 수고가 작았다 (Fig. 2). 지수모델의 경우 시비 시작 8주차부터 시비량이 급격하게 증가하였고 전체 시비량은 관행 시비량의 2배 (8주차, 25 vs 45 (관행 vs 지수) mg seedling-1; 9주차, 25 vs 65 mg seedling-1)였다 (Fig. 1). 육묘 중반 이후 묘목 성장 속도에 따른 질소 요구량을 상회하여 시비량을 급격하게 증가시켰기 때문에 뿌리 내 역삼투압에 의해 수분 흡수 불량 및 묘목 고사로 인해 (Albornoz, 2016) 9주 차 이후 지수모델 처리구에서의 느티나무 성장이 저해된 것으로 판단된다. 느티나무는 해송 (내염성 강)과 상수리 (내염성 중)에 비해 내염성이 낮기 때문 (Ahn et al., 2008)으로 판단된다. 따라서, 수종 별 내염성을 반영하여 시비량을 결정할 필요가 있을 것으로 판단된다.
식물체 부위별 건물중은 수고와 양상이 달랐다 (Fig. 2). 묘목 수고는 시비 처리에 따른 유의한 차이가 없는 반면, 건물중은 해송과 상수리의 경우 관행과 수정지수 모델의 건물중이 지수모델 시비 처리구보다 유의하게 높았다. 수고 차이는 없지만 건물중이 더 큰 이유는 관행과 수정지수 시비 처리에 의해 길이 생장 보다는 너비 또는 부피 생장이 더 잘 일어났음을 의미한다. 느티나무의 경우 수정지수 모델의 건물중이 관행과 지수모델 시비 처리구보다 더 컸는데, 지수모델의 경우 육묘 9 주 이후 성장이 저해된 결과로 판단된다. 관행과 수정지수 모델을 비교하였을 때, 두 처리에서 수고의 차이는 없었지만, 수정지수 처리구에서 건물중이 유의하게 증가하였는데, 이는 관행에 비해 수정지수 모델에서 너비 또는 부피 성장이 우수하였기 때문으로 판단된다. 세 수종 모두 지수모델로 시비를 하였을 때 관행과 수정지수모델에 비해 건물중이 작았는데, 이는 성장 초기 양분이 부족하여 성장이 충분하지 않았으며 생육 중반 이후 급격한 시비량 증가에 따른 독성에 의해 성장이 저해되었기 때문으로 판단된다 (Hu, 2012).
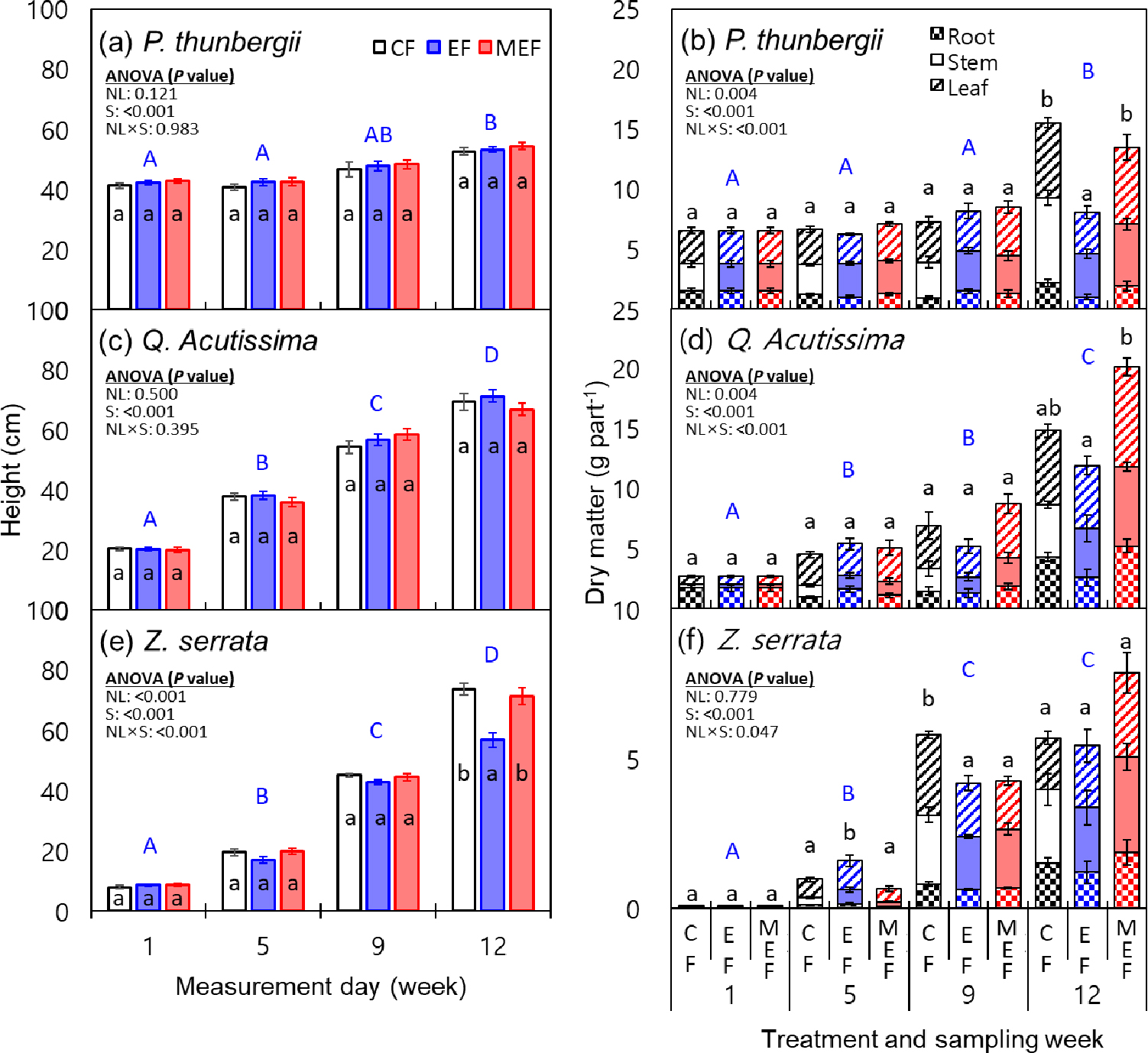
Fig. 2.
Height (a, c, and e) and dry matter (b, d, and f) of tree seedlings grown by different nutrient loading models; CF, conventional fertilization model; EF, exponential fertilization model; MEF, modified exponential model. Same lowercase letters are not significantly different at α = 0.05 among the nutrient loading models. Same uppercase letters are not significantly different at α = 0.05 among the measurement (sampling) days. ANOVA legends; NL, nutrient loading; S, sampling period.
시비 처리에 따른 묘목 내 양분함량 변화
수종 별 시비 처리에 따른 탄소 (mg C part-1)와 질소 (mg N part-1) 함량 변화는 묘목 건물중 변화와 유사하였다 (Fig. 3). 식물체내 부위별 탄소와 질소 농도는 부위 별 차이는 있었지만, 동일 부위의 경우 시비 처리에 따른 차이는 없었다. 따라서 부위 별 질소 함량은 건물중에 의해 결정되었다. Salifu and Timmer (2003)에 따르면 시비량 증가에 따른 묘목 내 양분 함량이 부족 (deficiency) - 과잉흡수 (luxury consumption) - 독성 (toxicity) 단계로 변화하는데, 과잉흡수 상태에 있을 때 양분집적이 되었다고 판단한다. 양분 부족 상태와 과잉흡수 상태 사이는 식물체 건물중은 비슷하지만 양분 농도 또는 함량이 과잉 흡수 상태에서 더 높게 나타난다. 따라서 본 연구에서 사용한 지수모델 또는 수정 지수모델에 의해 묘목 내 질소 함량이 관행모델에 비해 증가한 것은 지수모델이나 수정지수모델에 의해 양분이 집적되었음을 의미한다.
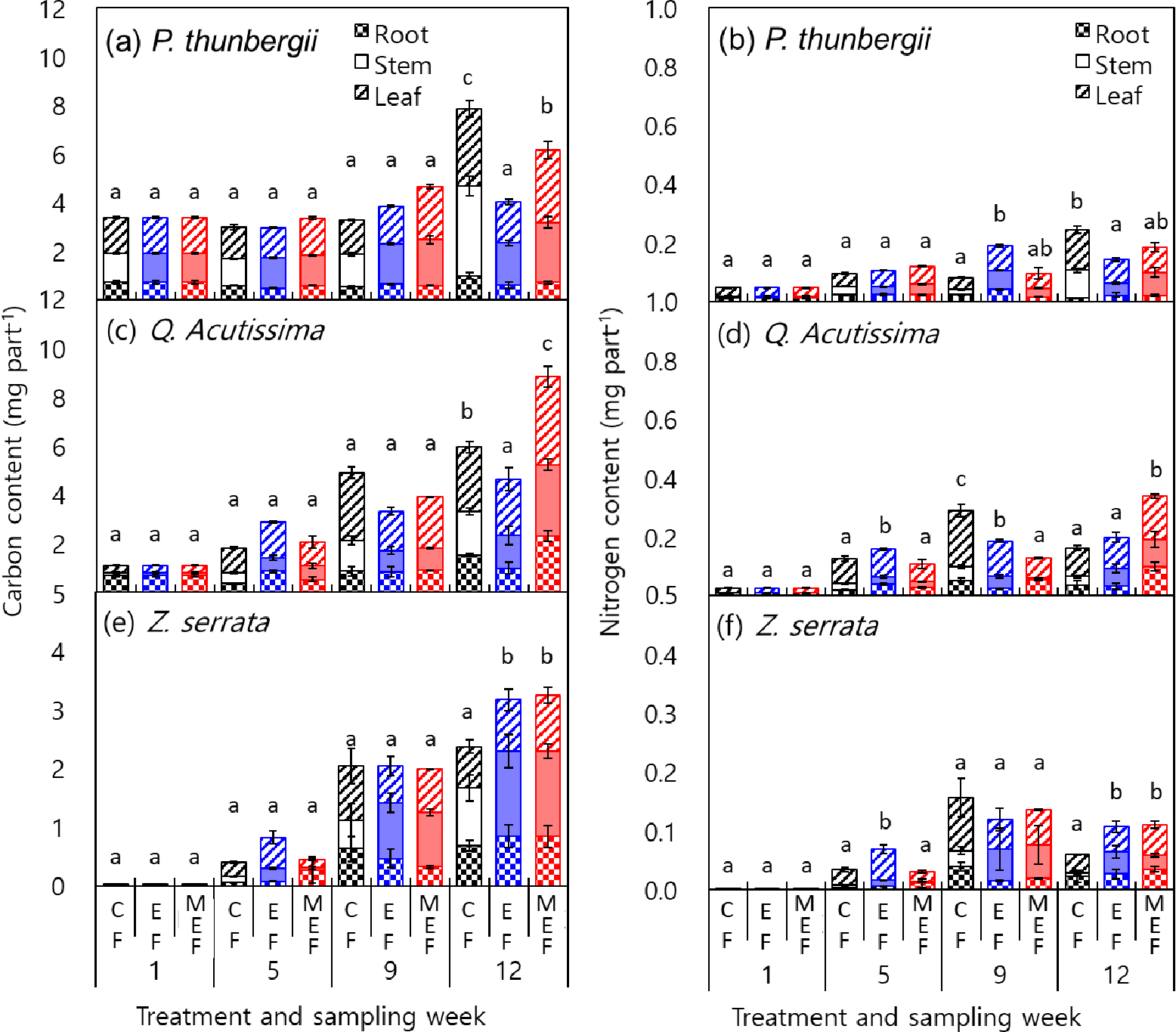
Fig. 3.
Carbon content (a, c, and e) and nitrogen content (b, d, and f) of each part of tree seedlings grown by different nutrient loading models; CF, conventional fertilization model; EF, exponential fertilization model; MEF, modified exponential model. Same lowercase letters are not significantly different at α = 0.05 among the nutrient loading models.
관련 연구 (Hu, 2012; Pokharel et al., 2017)에 따르면 방크스소나무 (jack pine, Pinus banksiana), 글라우카가문비나무 (white spruce, Picea glauca), 사시나무 (trembling aspen, Populus tremuloides) 수종에 대해 양분집적 효율을 평가한 결과, 수정지수 모델이 양분집적에 가장 효율적이었으며, 양분집적된 묘목이 관행 육묘 묘목에 비해 토양 양분 함량이 낮고 잡초와의 양분경쟁이 심한 열악한 조건에서도 현장 생존율과 성장량이 우수하다고 평가되었다. 본 연구에서도 관행모델에 비해 수정지수모델이 양분집적에 더 효율적인 것으로 판단되었다. 하지만 지수모델에 의해 생육 중반 이후 과잉 시비에 의해 묘목이 고사하는 현상이 나타났으며, 이를 해결하기 위해서는 육묘 기간 중 총 시비량을 다르게 설정 후 생육 시기별 시비량 조절이 필요할 것으로 판단된다.
최적의 양분집적 묘목은 묘목의 크기는 관행에 비해 크지 않지만 식물체내 양분 함량이 관행에 비해 높은 것이다 (Imo and Timmer, 2002). 식물체내 양분함량이 높으면 간척지나 광산 채굴 후 복원된 지역과 같이 토양 물리화학적 조건이 식물이 자라기 부적합한 조건에서도 묘목 이식 후 토양 적응성이 우수하고 잡초와의 양분 경쟁에서 우수하기 때문에 (Pokharel et al., 2017) 묘목 이식에 필요한 토양 개량제, 비료, 제초제 또는 잡초매트 등의 비용 절감에 효과적일 것으로 판단된다.
Conclusions
국내 대표 수종인 해송, 느티나무, 상수리에 대하여 유묘의 양분 집적 가능 여부에 대하여 연구를 수행하였다. 양분 집적 모델과 상관없이 묘목 수고는 차이가 없었으나 건물중은 관행 시비에 비해 수정 지수모형에서 더 높았다. 12주 후 묘목 부위별 질소 함량 또한 수정 지수모형에서 다른 모형에 비해 높아서 수정 지수 모형의 양분집적 효율이 가장 높음을 알 수 있다. 이는 수정 지수 모형은 생육 초기 양분을 보상해주고 생육 후기 양분 공급량이 관행에 비해 높기 때문으로 판단된다. 지수 모형의 경우 급격한 시비량 증가에 따라 고사 비율이 높았는데, 향후 연구에서는 다양한 시비량에 대해 평가를 하고, 간척지와 같은 현장 실험을 통해 양분 집적 묘목의 노지 적응성 및 성장에 대한 평가가 필요할 것으로 판단된다.
